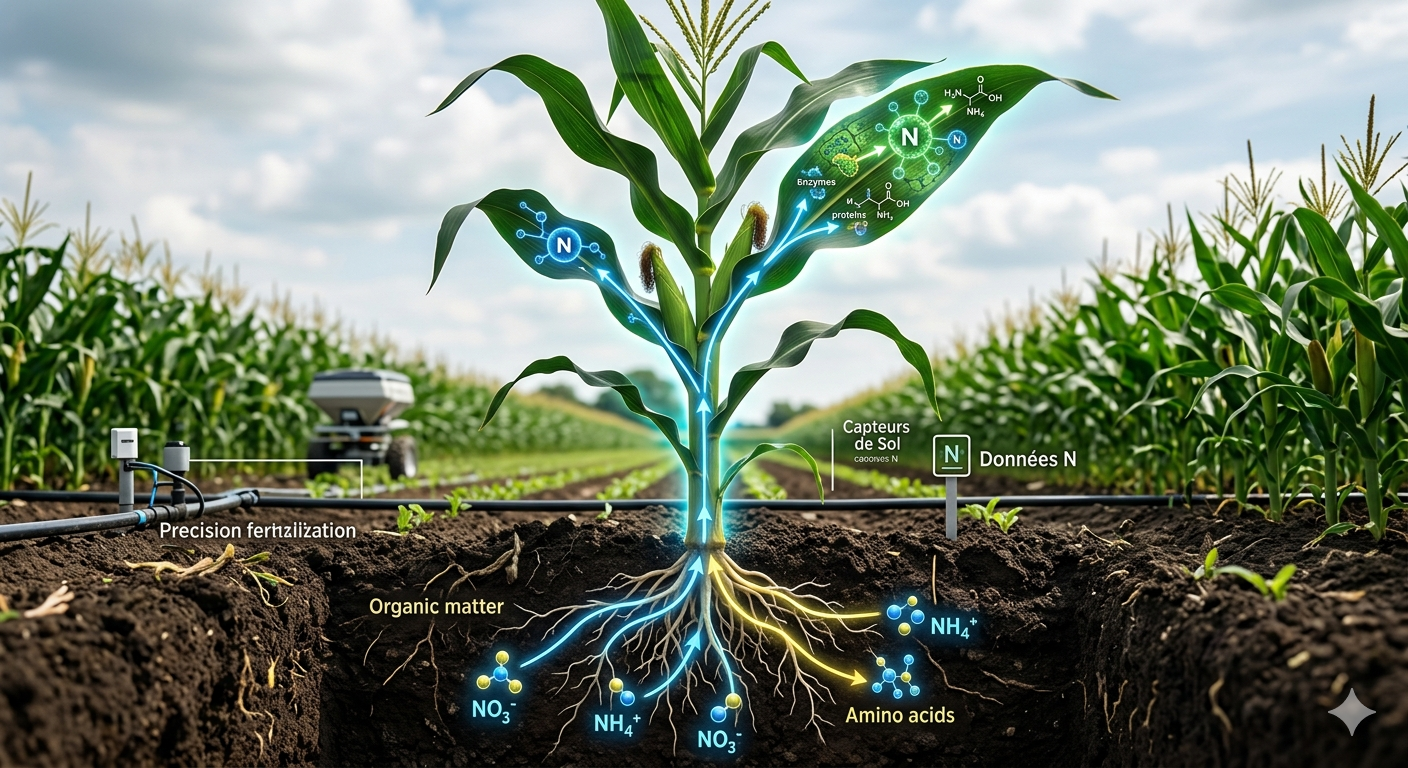
Résumé : L'azote est l'élément nutritif le plus limitant pour la croissance du maïs, et son assimilation conditionne directement le rendement et la qualité des grains. Le maïs absorbe l'azote sous deux formes principales : les nitrates (NO₃⁻) et l'ammonium (NH₄⁺), présents dans la solution du sol à l'issue du cycle biogéochimique de l'azote. La disponibilité relative de ces deux formes dépend de l'activité microbienne (nitrification), des conditions physico-chimiques du sol (pH, aération, température) et des pratiques de fertilisation. Le maïs possède des systèmes de transport spécifiques pour chaque forme : les transporteurs de nitrates (NRT) et les transporteurs d'ammonium (AMT), dont l'expression est finement régulée par les besoins de la plante et la disponibilité du sol. L'assimilation de l'azote en nitrates et ammonium implique des voies métaboliques distinctes mais complémentaires, avec des coûts énergétiques différents qui influencent l'efficience d'utilisation de l'azote. La compréhension de ces mécanismes est fondamentale pour optimiser les pratiques de fertilisation : fractionnement des apports, choix des formes d'engrais (ammonitrates, urée, engrais organiques), utilisation de produits inhibiteurs de nitrification, et sélection de variétés à meilleure efficience. Cette connaissance permet de réduire les pertes d'azote vers l'environnement (lessivage des nitrates, émissions de N₂O) tout en maintenant la productivité.
1. Le cycle de l'azote dans le sol : des formes minérales disponibles pour le maïs
1.1 Les transformations de l'azote dans le sol
L'azote présent dans le sol se trouve sous deux grandes catégories de formes. D'une part, l'azote organique, contenu dans la matière organique du sol, les résidus de culture et les effluents d'élevage, qui n'est pas directement assimilable par les plantes. D'autre part, l'azote minéral, qui comprend les nitrates (NO₃⁻) et l'ammonium (NH₄⁺), et qui constitue la forme directement assimilable par le maïs.
Le passage de l'azote organique à l'azote minéral est assuré par l'activité des micro-organismes du sol via le processus de minéralisation. Ce processus biologique est influencé par plusieurs facteurs environnementaux. La température joue un rôle clé, avec une activité optimale entre 25 et 35°C. L'humidité du sol est également déterminante : l'activité microbienne est maximale lorsque le sol est à capacité au champ. Le pH du sol influence l'activité des micro-organismes, avec des conditions neutres les plus favorables. Enfin, la texture et la teneur en matière organique du sol conditionnent la capacité de minéralisation.
1.2 La nitrification : transformation de l'ammonium en nitrate
L'ammonium (NH₄⁺) issu de la minéralisation ou des apports d'engrais est transformé en nitrates (NO₃⁻) par un processus appelé nitrification, réalisé par des bactéries spécifiques. Ce processus se déroule en deux étapes successives.
La première étape, la nitritation, est réalisée par des bactéries du genre Nitrosomonas et d'autres espèces apparentées. Ces micro-organismes oxydent l'ammonium (NH₄⁺) en nitrites (NO₂⁻). La seconde étape, la nitratation, est assurée par des bactéries du genre Nitrobacter et d'autres espèces, qui oxydent les nitrites (NO₂⁻) en nitrates (NO₃⁻).
Cette transformation a des conséquences majeures pour la gestion de la fertilisation azotée. Les nitrates sont très mobiles dans le sol car ils ne sont pas retenus par le complexe argilo-humique (ce sont des anions). Ils sont donc facilement lessivables par les pluies ou l'irrigation, ce qui peut entraîner des pertes d'azote et une pollution des eaux souterraines. En revanche, l'ammonium, étant un cation, est retenu sur le complexe argilo-humique et donc moins sujet au lessivage. Cependant, l'ammonium présent en surface peut être perdu par volatilisation ammoniacale, c'est-à-dire la transformation en ammoniac (NH₃) gazeux qui s'échappe dans l'atmosphère, particulièrement lorsque le pH du sol est élevé et que les températures sont chaudes.
1.3 Les pertes d'azote : des enjeux agronomiques et environnementaux majeurs
Les pertes d'azote du système sol-plante représentent une double perte. Sur le plan agronomique, elles se traduisent par une diminution de l'efficacité de la fertilisation et une baisse du rendement potentiel. Sur le plan environnemental, elles contribuent à la pollution des eaux souterraines par les nitrates et aux émissions de gaz à effet de serre, notamment le protoxyde d'azote (N₂O).
Plusieurs mécanismes de perte sont à distinguer. Le lessivage concerne principalement les nitrates. Il se produit lorsqu'il y a un excès d'eau qui entraîne les nitrates en profondeur, hors de la zone d'absorption racinaire. Ce phénomène est particulièrement marqué sur les sols sableux et lorsque les apports d'azote sont réalisés hors période de prélèvement actif par la culture.
La dénitrification est un processus biologique par lequel les nitrates sont transformés en azote gazeux (N₂) et en protoxyde d'azote (N₂O). Ce phénomène se produit en conditions de sol saturé en eau, où l'oxygène est limité, et en présence de carbone organique qui sert de substrat énergétique aux bactéries dénitrifiantes.
La volatilisation ammoniacale concerne l'ammonium apporté en surface. Lorsque l'ammonium est exposé à l'air et que le pH du sol est élevé, une partie se transforme en ammoniac gazeux (NH₃) et s'échappe dans l'atmosphère. Ce phénomène est particulièrement important lors des apports d'urée non enfouis, surtout par temps chaud et venteux.
Enfin, l'immobilisation correspond à la transformation de l'azote minéral en azote organique par les micro-organismes du sol. Ce phénomène se produit lorsque les résidus de culture apportés ont un rapport carbone sur azote (C/N) élevé, typiquement avec les pailles de céréales. Les micro-organismes puisent alors l'azote minéral du sol pour décomposer ces résidus, le rendant temporairement indisponible pour la culture.
2. Les systèmes de transport de l'azote chez le maïs
2.1 Une adaptation à deux formes minérales
Le maïs, comme la plupart des plantes supérieures, a développé des systèmes de transport spécifiques pour capturer l'azote du sol sous ses deux formes minérales. Ces systèmes sont localisés principalement au niveau des racines, et leur expression est finement régulée à la fois par les besoins de la plante et par la disponibilité de l'azote dans la solution du sol. Cette double régulation permet d'adapter l'absorption aux conditions changeantes du milieu tout en évitant une absorption excessive qui pourrait être toxique.
2.2 Les transporteurs de nitrates (NRT)
Les transporteurs de nitrates, abrégés NRT, sont des protéines membranaires qui assurent l'entrée du NO₃⁻ dans les cellules racinaires. On distingue deux grandes familles de transporteurs aux propriétés différentes.
La première famille, appelée NRT1 (ou NPF), regroupe des transporteurs de faible affinité mais de haute capacité. Ces transporteurs fonctionnent lorsque la concentration en nitrates dans le sol est élevée, typiquement après un apport d'engrais. Ils assurent un transport massif de nitrates et jouent également un rôle dans le chargement de ces nitrates dans le xylème pour leur transport vers les parties aériennes.
La seconde famille, appelée NRT2, comprend des transporteurs de haute affinité mais de faible capacité. Ils sont particulièrement importants lorsque la concentration en nitrates dans le sol est faible, en situation de carence. Leur expression est fortement induite par les conditions de carence azotée. Chez le maïs, le gène ZmNRT2.1 est un exemple typique de transporteur de haute affinité : il est fortement induit en conditions de carence azotée, permettant à la plante d'augmenter sa capacité d'absorption lorsque les nitrates se raréfient.
La famille NPF (anciennement NRT1) chez le maïs est particulièrement diversifiée et comprend de nombreux membres aux fonctions variées. Au-delà du transport des nitrates, certains de ces transporteurs interviennent dans le transport d'autres substrats comme les acides aminés, les peptides, ou encore les hormones végétales, illustrant la multifonctionnalité de cette famille de protéines.
2.3 Les transporteurs d'ammonium (AMT)
Les transporteurs d'ammonium, abrégés AMT, assurent l'entrée du NH₄⁺ dans les cellules racinaires. Comme pour les nitrates, on distingue plusieurs familles aux propriétés complémentaires.
La famille AMT1 regroupe des transporteurs de haute affinité, dont l'expression est induite par les conditions de carence azotée. Ils permettent à la plante de capter efficacement l'ammonium lorsque sa concentration dans le sol est faible.
La famille AMT2 comprend des transporteurs de faible affinité, dont l'expression est souvent constitutive, c'est-à-dire maintenue à un niveau de base indépendamment des conditions de disponibilité.
L'assimilation directe de l'ammonium présente un avantage énergétique par rapport aux nitrates, car l'ammonium est déjà à l'état réduit alors que les nitrates doivent subir une réduction avant d'être incorporés dans les acides aminés. En revanche, l'ammonium en forte concentration peut être toxique pour les plantes, provoquant des désordres métaboliques et une acidification du cytoplasme. Cette toxicité potentielle explique l'existence de mécanismes de régulation fine de l'absorption de l'ammonium, empêchant une accumulation excessive dans les cellules.
2.4 La régulation de l'absorption : une adaptation aux besoins
L'absorption de l'azote par le maïs n'est pas un processus passif ; elle est finement régulée en fonction des besoins de la plante et de la disponibilité du sol. Plusieurs mécanismes de régulation sont impliqués à différents niveaux.
La régulation transcriptionnelle correspond à l'induction ou à la répression de l'expression des gènes codant pour les transporteurs. Par exemple, en situation de carence azotée, la plante active la transcription des gènes NRT2 et AMT1 pour augmenter sa capacité d'absorption. Inversement, en situation d'abondance, l'expression de ces gènes est réprimée pour éviter une absorption excessive.
La régulation post-traductionnelle agit directement sur les protéines transporteurs déjà synthétisées. En conditions de forte disponibilité en azote, certaines protéines transporteurs peuvent être dégradées ou inactivées, permettant une réponse rapide sans attendre la modification de l'expression des gènes.
Enfin, une signalisation systémique relie les besoins de la partie aérienne aux racines. Lorsque les feuilles sont déficitaires en azote, elles émettent des signaux (peptides, hormones) qui sont transportés vers les racines et modulent l'expression des transporteurs. Cette communication à distance permet d'ajuster précisément l'absorption aux besoins réels de la plante entière.
3. Les voies d'assimilation : de l'azote minéral aux acides aminés
3.1 L'assimilation des nitrates
Les nitrates absorbés par les racines peuvent suivre trois destinées. Une partie peut être stockée dans les vacuoles des cellules racinaires, constituant un réservoir temporaire. Une autre partie est transportée vers les parties aériennes via le xylème, pour être assimilée dans les feuilles. Enfin, une fraction peut être réduite directement dans les racines.
La réduction des nitrates en ammonium est une voie métabolique en deux étapes, catalysée par deux enzymes successives. La première étape est réalisée par la nitrate réductase (NR), une enzyme localisée dans le cytosol. Elle réduit les nitrates (NO₃⁻) en nitrites (NO₂⁻) en utilisant l'énergie fournie par le NAD(P)H. La seconde étape est catalysée par la nitrite réductase (NiR), localisée dans les chloroplastes des feuilles ou dans les plastes des racines. Cette enzyme réduit les nitrites (NO₂⁻) en ammonium (NH₄⁺). La réduction des nitrates est énergétiquement coûteuse : elle nécessite deux équivalents de NAD(P)H par nitrate réduit, ce qui explique pourquoi l'assimilation des nitrates a un coût énergétique supérieur à celle de l'ammonium.
3.2 L'assimilation de l'ammonium
L'ammonium, qu'il provienne directement de l'absorption racinaire ou de la réduction des nitrates, est rapidement incorporé dans les acides aminés via un cycle métabolique appelé cycle GS/GOGAT, qui constitue la porte d'entrée de l'azote dans le métabolisme carboné.
La première étape est catalysée par la glutamine synthétase (GS). Cette enzyme combine l'ammonium (NH₄⁺) avec du glutamate, en présence d'ATP (énergie), pour former de la glutamine. La seconde étape est réalisée par la glutamate synthase (GOGAT), qui transfère le groupe amine de la glutamine vers l'α-cétoglutarate (un acide organique issu du cycle de Krebs) pour former deux molécules de glutamate. L'une de ces deux molécules de glutamate peut être réutilisée par la glutamine synthétase, fermant ainsi le cycle.
Le glutamate ainsi formé sert de donneur d'azote pour la synthèse des autres acides aminés. Par l'action d'enzymes appelées transaminases, le groupe amine du glutamate est transféré à différents squelettes carbonés (acides α-cétoniques) pour produire l'ensemble des acides aminés nécessaires à la plante.
3.3 La formation des protéines et la croissance
Les acides aminés issus du cycle GS/GOGAT sont ensuite utilisés pour la synthèse des protéines dans les ribosomes des cellules, par le processus de traduction. Ils sont également transportés via le phloème vers les organes en croissance (jeunes feuilles, tiges, épis en formation) où ils sont utilisés pour la construction de nouveaux tissus. En fin de cycle, lors du remplissage du grain, les acides aminés sont massivement remobilisés des feuilles et des tiges vers l'épi, où ils sont incorporés dans les grains sous forme de protéines de réserve.
Chez le maïs, les protéines de réserve des grains sont principalement des zéines. La teneur en protéines des grains de maïs, généralement comprise entre 7 et 10 %, est directement liée à la disponibilité en azote pendant la période critique du remplissage. Une carence azotée à ce stade se traduit par une baisse du rendement (moins de grains ou grains plus légers), une diminution de la teneur en protéines, et une moindre valeur alimentaire pour l'élevage, particulièrement pour les monogastriques (porcs, volailles) qui ont des besoins en acides aminés spécifiques.
4. Efficience d'utilisation de l'azote chez le maïs
4.1 Définition et enjeux
L'efficience d'utilisation de l'azote, abrégée NUE (Nitrogen Use Efficiency), est le rapport entre le rendement en grain (exprimé en kilogrammes) et la quantité d'azote disponible pour la plante (azote du sol apporté par la minéralisation et azote apporté par les engrais). Une NUE élevée signifie que la plante produit un maximum de grain avec un minimum d'azote, ce qui réduit d'autant les pertes vers l'environnement.
La NUE peut être décomposée en deux composantes principales. L'efficience d'absorption correspond à la capacité de la plante à prélever l'azote du sol. Elle dépend du développement du système racinaire (profondeur, densité, ramification), de l'expression des transporteurs d'azote, et des interactions avec les micro-organismes du sol (mycorhizes notamment).
L'efficience d'utilisation correspond à la capacité de la plante à convertir l'azote absorbé en rendement de grain. Elle dépend de l'allocation de l'azote entre les différents organes, de l'efficacité du métabolisme, et de la capacité à remobiliser l'azote des parties aériennes vers les grains en fin de cycle.
4.2 Variabilité génétique de l'efficience azotée
Il existe une variabilité génétique naturelle de l'efficience d'utilisation de l'azote chez le maïs, ce qui ouvre la voie à des programmes de sélection variétale. Les objectifs de sélection sont multiples.
L'objectif de développer des variétés plus économes en azote vise à maintenir un rendement élevé avec des apports d'engrais réduits. Ces variétés sont particulièrement adaptées aux systèmes à bas intrants et aux contextes où le coût des engrais est élevé.
L'objectif d'améliorer l'efficience d'absorption cible des traits racinaires spécifiques : systèmes racinaires plus profonds pour capter l'azote lessivé en profondeur, ou plus denses pour explorer plus finement le volume de sol.
L'objectif d'améliorer la remobilisation consiste à sélectionner des variétés capables de transférer efficacement l'azote des feuilles et des tiges vers les grains en fin de cycle, ce qui est particulièrement important pour les variétés à cycle long.
4.3 Le rôle de la date de floraison
La période autour de la floraison est critique pour l'absorption et l'allocation de l'azote chez le maïs. C'est à ce moment que se met en place le nombre potentiel de grains (détermination du nombre d'ovules fécondés), et que commence le remplissage des grains. Un stress azoté à ce stade a des conséquences majeures sur le rendement final, car il limite à la fois le nombre de grains fixés et leur poids final.
Les variétés de maïs ont été sélectionnées pour synchroniser la demande en azote du grain avec la capacité de fourniture du sol et de l'engrais. La durée de la période de remplissage est un caractère important : des variétés à remplissage long peuvent mieux valoriser des apports fractionnés en fin de cycle, car elles maintiennent une activité photosynthétique et une capacité d'absorption plus longtemps.
5. Optimisation de la fertilisation azotée pour le maïs
5.1 Diagnostic des besoins
Avant tout apport d'engrais, un diagnostic précis permet d'ajuster les doses aux besoins réels de la culture, évitant à la fois le gaspillage et les carences.
L'analyse de terre est la méthode de base. Elle mesure l'azote minéral présent dans le sol (le reliquat sortie hiver), c'est-à-dire l'azote qui sera disponible pour la culture en début de cycle. Cette mesure permet de calculer la dose d'engrais à apporter en fonction de l'objectif de rendement et du type de sol.
L'analyse de plante, réalisée sur les feuilles à différents stades, renseigne sur la nutrition réelle de la culture. La teneur en azote des feuilles peut être comparée à des seuils de référence pour ajuster les apports de couverture.
Les indices de végétation, mesurés par capteurs embarqués sur tracteurs, par drones ou par satellites, permettent de cartographier les zones carencées au sein d'une même parcelle. L'agriculture de précision utilise ces informations pour moduler les doses d'engrais en temps réel.
Les modèles de simulation intègrent les données climatiques, les caractéristiques du sol, et les paramètres de la culture pour simuler la dynamique de l'azote et proposer des scénarios d'optimisation.
5.2 Choix des formes d'engrais
Le choix de la forme d'engrais azoté influence la disponibilité de l'azote pour la plante et les risques de pertes.
Les ammonitrates (engrais contenant à la fois de l'ammonium et des nitrates) présentent l'avantage de fournir les deux formes assimilables immédiatement. Leur disponibilité est rapide, mais la fraction nitrate est sensible au lessivage. Le fractionnement des apports est recommandé pour limiter ce risque.
L'urée est un engrais très concentré (46 % d'azote) et généralement moins coûteux. Cependant, l'urée doit être transformée en ammonium par une enzyme (uréase) présente dans le sol avant d'être assimilable. Cette transformation s'accompagne d'un risque de volatilisation ammoniacale si l'urée n'est pas rapidement enfouie. Des inhibiteurs d'uréase peuvent être utilisés pour réduire ce risque.
Les engrais ammoniacaux (sous forme d'ammonium) sont retenus sur le complexe argilo-humique et donc moins lessivables. Cependant, ils subissent progressivement une nitrification qui les transforme en nitrates. Ils sont particulièrement adaptés aux sols filtrants où le lessivage est un risque majeur.
Les engrais organiques (fumiers, lisiers, composts, digestats de méthanisation) apportent à la fois de l'azote et de la matière organique, qui améliore la structure et la fertilité du sol. Leur azote est libéré progressivement par minéralisation, ce qui réduit les risques de perte mais rend la disponibilité moins prévisible. Leur utilisation est particulièrement pertinente dans les sols à faible teneur en matière organique.
5.3 Le fractionnement : une pratique clé
Le fractionnement des apports d'azote consiste à répartir la dose totale en plusieurs fractions appliquées à différents stades de la culture. Cette pratique permet d'aligner la disponibilité de l'azote avec les besoins réels de la plante, réduisant ainsi les périodes pendant lesquelles l'azote est présent dans le sol sans être utilisé par la culture, et donc exposé aux pertes.
L'apport de fond (pré-semis) consiste à apporter une partie de l'azote, souvent sous forme ammoniacale, avant le semis. Cette fraction assure le démarrage de la culture et peut être localisée pour limiter les pertes.
L'apport au semis, appelé "starter", est une petite dose d'azote localisée au niveau de la ligne de semis. Il favorise l'implantation rapide des jeunes plantes, particulièrement en conditions de sol froid ou en semis direct.
Les apports de couverture sont réalisés en cours de végétation, généralement au stade 4 à 6 feuilles, et parfois également avant la floraison. Ces fractions permettent d'ajuster la nutrition en fonction du développement réel de la culture et des conditions météorologiques.
Le fractionnement permet de réduire les doses totales d'azote à rendement équivalent, car moins d'azote est perdu avant d'être absorbé par la plante.
5.4 Les inhibiteurs de nitrification
Les inhibiteurs de nitrification (IN) sont des produits qui ralentissent l'activité des bactéries responsables de la transformation de l'ammonium en nitrate. Leur intérêt est multiple.
En maintenant l'azote sous forme ammonium, moins lessivable, les inhibiteurs réduisent le risque de lessivage, particulièrement sur les sols filtrants ou en cas de pluies précoces.
Le maïs préfère souvent une alimentation mixte (ammonium + nitrate) plutôt qu'une alimentation exclusivement nitrique. Le maintien d'une proportion d'ammonium peut favoriser la croissance.
Les émissions de protoxyde d'azote (N₂O), un puissant gaz à effet de serre, sont produites lors de la nitrification et de la dénitrification. En ralentissant la nitrification, les inhibiteurs réduisent ces émissions.
Des produits comme la dicyandiamide (DCD) ou le nitrapyrine sont utilisés en complément des engrais azotés, notamment pour les apports précoces sur sols filtrants ou en conditions climatiques favorables aux pertes.
5.5 L'intégration dans des systèmes durables
La gestion de l'azote pour le maïs s'inscrit dans des systèmes de culture plus larges, où plusieurs pratiques interagissent.
Les cultures intermédiaires (CIPAN) piègent l'azote résiduel après la récolte, réduisant le lessivage hivernal et restituant cet azote à la culture suivante.
La rotation culturale permet d'introduire des légumineuses (pois, féverole, luzerne) qui fixent l'azote atmosphérique, réduisant les besoins en engrais pour la culture suivante.
Les apports organiques (fumiers, composts, digestats) valorisent les effluents d'élevage et les coproduits, bouclant les cycles de matière à l'échelle de l'exploitation ou du territoire.
Les systèmes sans labour (semis direct, travail superficiel) préservent la matière organique du sol et l'activité biologique, favorisant une minéralisation progressive de l'azote mieux synchronisée avec les besoins de la culture.
6. Recommandations pratiques pour la fertilisation du maïs
6.1 Pour les agriculteurs
Plusieurs pratiques permettent d'optimiser la gestion de l'azote. La réalisation d'un reliquat sortie hiver est essentielle pour connaître l'azote minéral disponible dans le sol avant le démarrage de la culture. Cette analyse constitue la base du raisonnement de la fertilisation.
Le fractionnement des apports en deux ou trois fractions, selon le type de sol et les prévisions météorologiques, permet de synchroniser la disponibilité de l'azote avec les besoins de la culture.
La localisation de l'azote starter au semis, en petite dose au niveau de la ligne de semis, favorise l'implantation et permet de réduire les apports de fond.
L'utilisation d'inhibiteurs de nitrification est particulièrement pertinente sur les sols filtrants ou pour les apports précoces, afin de réduire les risques de lessivage.
L'adaptation des doses à la variabilité intra-parcellaire, via l'agriculture de précision (capteurs embarqués, cartes de rendement), permet d'ajuster les apports aux zones carencées et de réduire les apports là où la disponibilité naturelle est suffisante.
Enfin, l'intégration de la fertilisation azotée dans la rotation est essentielle. La valeur de l'azote fixé par les légumineuses précédentes doit être prise en compte, et les doses d'engrais doivent être adaptées en conséquence.
6.2 Pour les conseillers
Les conseillers agricoles ont un rôle clé à jouer dans l'accompagnement des agriculteurs vers une gestion optimisée de l'azote. La formation aux nouvelles techniques de diagnostic, notamment l'interprétation des reliquats et l'utilisation des capteurs, est indispensable.
Le développement de références locales pour l'interprétation des analyses permet d'ajuster les recommandations aux conditions pédoclimatiques spécifiques de chaque région.
La promotion du fractionnement comme levier de réduction des pertes doit s'appuyer sur des démonstrations et des références technico-économiques locales.
L'accompagnement à l'utilisation des inhibiteurs de nitrification doit se faire dans les contextes où leur efficacité est avérée, en tenant compte des conditions de sol et des pratiques culturales.
6.3 Pour les pouvoirs publics et la recherche
Le soutien à la recherche sur la variabilité génétique de l'efficience azotée chez le maïs est essentiel pour développer des variétés plus économes. Les programmes de sélection doivent être encouragés et financés.
Le financement de la mise au point d'outils de diagnostic (capteurs de terrain, modèles de simulation, algorithmes d'IA) permet de rendre ces technologies accessibles aux agriculteurs.
L'encouragement des pratiques vertueuses via les aides de la Politique Agricole Commune, notamment les écorégimes et les mesures agro-environnementales, contribue à la diffusion des bonnes pratiques.
Conclusion
La compréhension des mécanismes d'assimilation de l'azote par le maïs a considérablement progressé ces dernières décennies, permettant aujourd'hui d'optimiser la fertilisation avec une précision inégalée.
Le maïs absorbe l'azote sous deux formes, les nitrates et l'ammonium, via des transporteurs spécifiques dont l'expression est finement régulée par les besoins de la plante. L'assimilation de ces deux formes mobilise des voies métaboliques distinctes, avec un coût énergétique plus élevé pour les nitrates que pour l'ammonium.
L'efficience d'utilisation de l'azote est un caractère complexe, mais il existe une variabilité génétique naturelle qui peut être exploitée par la sélection pour développer des variétés plus économes et plus adaptées aux systèmes à bas intrants.
Les pratiques de fertilisation ont un impact majeur sur l'efficience et les pertes. Le fractionnement des apports, le choix des formes d'engrais, l'utilisation d'inhibiteurs de nitrification, et l'intégration dans des systèmes de culture diversifiés (rotations, cultures intermédiaires, apports organiques) sont autant de leviers pour améliorer la performance agronomique et environnementale.
Les enjeux sont considérables. L'azote est le premier poste de consommation d'énergie en agriculture, du fait de la fabrication des engrais par le procédé Haber-Bosch, très énergivore. Ses pertes vers l'environnement (lessivage des nitrates dans les eaux souterraines, émissions de protoxyde d'azote dans l'atmosphère) sont une préoccupation majeure de santé publique et de climat.
Optimiser la nutrition azotée du maïs, c'est à la fois améliorer la rentabilité des exploitations, réduire l'empreinte carbone de la production, préserver la qualité des eaux, et atténuer le changement climatique. Pour l'avenir, la combinaison de variétés plus efficientes, d'outils de diagnostic précis (capteurs, imagerie, modèles) et de pratiques agronomiques raisonnées (fractionnement, inhibiteurs, intégration dans la rotation) permettra de concilier productivité et durabilité.